近期的研究显示静息心率与动物寿命呈负相关:在动物王国中,心率较快的恒温哺乳动物的寿命一般短于心率较慢的恒温哺乳动物;就单个恒温哺乳动物而言,例如人类,有几项在健康人、高血压患者以及冠心病患者中的大样本研究,提示较快的静息心率几乎普遍伴随更高的死亡风险。
 张卫国 美国德克萨斯州欧文市心血管和神经病研究所
张卫国 美国德克萨斯州欧文市心血管和神经病研究所
HR varies significantly among mammals: it can be as low as 30-35 beats per minute (bpm) in large animals like whales and elephants, or as high as 600-700 bpm in mice and even higher in shrews. Likewise, mammal lifespan also varies a great deal. Mammals that have slower average HRs tend to live much longer than those that have faster HRs. The relationship between HR and lifespan was well presented by Levine, who illustrated that HR is inversely correlated with lifespan in 15 mammal species. Although some variability inevitably exists, calculations using the available data based on observation yield a mean value of around 1x109 (one billion) heartbeats in a lifetime across almost all mammals (Figure 1).
A simplified way to approximately describe this phenomenon is that, on average, mammals have an allotment of 1x109 heart beats, and therefore those who use up their ultimate allotment faster will die sooner. Given a fact that the heart must contract continuously for many decades and cardiac muscle is unable to regenerate or repair itself, it is not unreasonable to assume that an animal is born with a heart which can only beat so many times before giving out.
In the same species, there are also dramatic variations in HR, and the same inverse correlations between average HR and lifespan. In humans, for example, several epidemiological investigations have shown that higher HR was strongly associated with cardiovascular mortality or sudden cardiac death. In a recent study of exercise and sudden death, HR was positively correlated with death from any cause, as well as sudden and non-sudden death from myocardial infarctions. In this study, the subjects were more stratified than in other studies, and even a difference of 15 bpm in resting HR almost doubled the risk of death from any cause and more than tripled the risk of sudden death from heart attack.
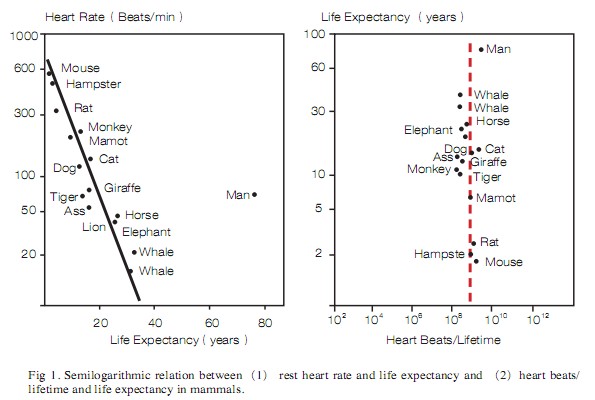
Are humans significantly different from other mammals in lifespan?
HR and lifespan are strongly correlated in mammals. Small mammals like mice, rats, and hamsters have the highest HRs and lowest life spans. Large mammals like elephants and whales have the slowest heat rates and the longest life spans. Humans, however, represent a distinct outlier in this relationship. Given an average human HR of 60–100 bpm, we should have similar life spans as tigers and giraffes, or about 25 years. As it is, however, our hearts can do over 3x109 (3 billion)beats in a lifetime. Why can humans achieve a significantly higher number of heart beats per lifetime than other mammals? Are humans physiologically or biologically different than other mammals in some way that allows us to “break” the relationship?
There are several caveats to the data point representing humans. For a majority of human history, average life span has been around 30 years. A life expectancy of 70-80 years in humans is mainly representative of developed nations lately. Acquisition of knowledge and development of technology in health care, disease prevention and treatment are primarily responsible for the increase in lifespan in these countries. In the US, for example, the life expectancy was reportedly below 50 years around 1900, but was close to 80 years in 2004. Given the fact that life expectancies of 40-50 years is still seen in some countries that lack adequate healthcare and disease control, it is clear that the distance between human lifespan and other mammals is largely attributable to acquired knowledge and skills in disease control and prevention, which has drastically lengthened life expectancy over last 1-2 centuries. In modern society, failures in the cardiovascular system are not necessarily fatal; modern medical practices such as cardiopulmonary resuscitation (CPR), defibrillation, and pacemaking prolong lives and save a great number of people who otherwise would have died.
Why is faster heart rate correlated with shorter lifespan in mammals?
Now the overarching questions are: why do smaller animals require faster HRs and why is faster HR so strongly correlated to shorter lifespan?
Higher resting HR results from higher basal metabolic rate. High metabolic rate is correlated with body surface area. Because smaller mammals have a greater ratio of body surface area to body weight, they need higher metabolic rates and more oxygen supply. This is essential for homeostasis in warm blooded mammals. According to scaling laws, body weight is a variable that determines HR. The relationship is written:HR oc BW1/4 or HR = C·BW1/4 .
Where HR is bpm, BW is body weight, and C is a coefficient. C varies slightly in the literature. Since HR has been shown to be correlated with lifespan, it follows that BW is correlated with lifespan as well. The average elephant lives 20 times as long as the average mouse because the HR in elephant is 20 times slower.
It is known that a faster HR is associated with the development of hypertension, atherosclerosis, sudden death and coronary heart disease, all leading to higher cardiovascular morbidity and mortality. In the general population, there is a 3-fold increase in mortality in subjects with HRs of 90-100 bpm compared to subjects with HRs of less than 60 bpm. In patients with hypertension, higher HR can lead to accelerated deterioration of the cardiovascular system. For example, in Framingham Study, increased HR is remarkably correlated with increased incidences cardiovascular and all cause mortality. In 2,293 elderly men and women, a 2-fold increase in mortality was seen in subjects with resting HR >79 bpm versus ≤79 bpm. Similarly, in the Coronary Artery Surgery Study, a 15 year follow-up of 24,913 patients with suspected or proven coronary disease, patients with resting HR 83 bpm at baseline had a significantly higher risk for total and cardiovascular mortality than those with HR <62 bpm.
In animal experiments, electrical pacing that increases HR is found to stiffen arteries, therefore causing progressive and profound reductions in arterial compliance and distensibility. At the molecular level, a 10%~20% increase in HR, independent of blood pressure or sympathetic nerve activity, significantly elevates cardiac NADPH oxidase activity and superoxide production, and activates mitogen-activated protein kinases, all indicating a greater oxidative stress to the heart and leading to cardiac hypertrophy and fibrosis. In dogs, increased reactive oxygen species (ROS)are considered to be an important component of the contractile dysfunction following rapid pacing. Previous studies indicate that the ROS generation rate in mitochondria plays a significant role in aging and longevity. Longevity is shortened by excessive generation of mitochondrial ROS that damages mitochondrial DNA and proteins.
High HR increases the expression of inflammation factors, such as interleukin-6 (IL-6), tumor necrosis factor alpha (TNF-α) and high sensitivity C-reactive protein(CRP). It has also been shown that the elevation of IL-6 and HR in heart failure patients can be simultaneously reduced by ??-blocker treatment. Some inflammatory cytokines can increase HR under experimental conditions. HR and biomarkers of inflammation are concurrently reduced by vagal nerve stimulation and by ??-adrenergic receptor blockade.
Faster HR often represents higher sympathetic nervous system activity. In some disease states, sympathetic activation is associa









 京公网安备 11010502033353号
京公网安备 11010502033353号